In processes like membrane trafficking, synaptic transmission, viral entry, infections or fertilization the regulation of membrane distances on a sub 10 nm-scale often plays a crucial role (Figure 1). However, it is not easy to determine membrane distances on such length scales. Classical tools to determine biological distances on sub 10 nm scales are based on Förster-energy transfer (FRET) type assays. Typically, the calculation of FRET distances rely on the presence of a single donor and a single acceptor. However, determining membrane distances is significantly more demanding since labeling the corresponding lipids of both membranes with donor and acceptor dyes intrinsically involves various different donor-acceptor molecule distances. This makes distance calculations from inter-membrane FRET signals difficult.
To be able to determine the distances of membranes without complex data analysis we have developed a calibrated membrane distance assay (Figure 2). To this end, we first tethered membranes of liposomes together at various distances by using DNA-strands of very well defined length. We then used the fluorescence lifetime of the donor as robust readout for the overall energy transfer efficiency observed at these distances. Using the observed values allows to generate calibration plots of donor fluorescence lifetimes versus membrane distances.

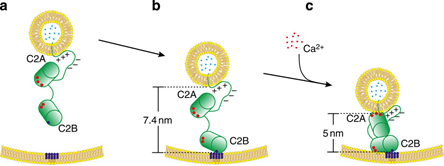
Applying this membrane distance ruler allows, for example, to investigate membrane distances hold by the synaptic protein Synaptotagmin 1 under different experimental conditions. For example, it has been proposed that one important function of the neuronal signal transmission trigger Synaptotagmin 1 is decreasing the membrane distance between synaptic vesicles containing the neurotransmitters and the presynaptic membrane upon an influx of Ca2+ as triggering signal. This could allow to reduce the membrane distance that enable the SNARE membrane fusion proteins to take action. The membrane fusion mediated by the SNARE proteins will then result ultimately in the release of the neurotransmitters into the synaptic cleft with subsequent transmitting the information to the postsynaptic nerve cell by neurotransmitter receptors. The results shown in figure provide evidence that Synaptotagmin 1 can indeed reduce membrane distances down to about 5 nm in the presence of Ca2+ at physiological triggering concentrations of ~100 µM. Careful control experiments were done to consider, for example, FRET to the DNA in the calibration experiments by measuring donor liposomes containing double strands of the calibration DNA in the absence of any acceptor liposomes.

These results provide the first direct experimental evidence that Synaptotagmin 1 can indeed regulate membrane distances on levels below 10 nm in the absence and presence of the synaptic transmission trigger Ca2+. In general, the membrane distance ruler developed in this project can be applied to a plethora of other mechanisms in which changes in membranes distances play an important role.
[1] C.-C. Lin, J. Seikowski, A. Pérez-Lara, R. Jahn, C. Höbartner, P. J. Walla, "Control of membrane gaps by synaptotagmin-Ca2+ measured with a novel membrane distance ruler", Nature Commu., 5, 5859 (2014).